🍉 受体蛋白
本页目收入有关overdose成瘾受体
本条目旨在从受体层面揭露耐药性,并提供在受体方面的解决办法
(部分见下文4、受体调节)一、什么是受体
受体(receptors),又称受器、接收器,是一个生物化学上的概念,
指一类能传导细胞外信号,并在细胞内产生特定效应的分子。
产生的效应可能仅在短时间内持续,比如改变细胞的代谢或者细胞的运动。
也可能是长效的效应,比如上调或下调某个或某些基因的表达。
受体通过与特定的配体结合而感知到细胞外的信号。随后,受体的结构发生变化,并诱导细胞内产生相应的效应。
受体通过信号级联应,逐步以指数级扩大细胞内产生的效应的强度。
信号级联的第一步可能是产生cAMP等第二信使分子,诱导下一级反应。
根据受体所在的置,可以分为细胞表面受体和细胞内受体两类。其中细胞表面受体位于细胞表面,处于内环境中的配体可以直接与之结合。
大部分的细胞内受都属于核受体。在未与配体结合时,这些受体位于细胞质中,配体需要穿过细胞膜进入细胞内,才能与该受体结合。
在与配体结合后,核受体会转入细胞核中发挥效应。另一类细胞内受体是细胞内的酶、RNA、核糖体等,
配体通过与这些受体结合发挥效应。
外部链接 维基百科
https://zh.wikipedia.org/wiki/受体_(生物化学)
https://zh.wikipedia.org/wiki/配體_(生物化學)
https://zh.wikipedia.org/wiki/核受体受体本身至少含有两个活性部位,一个是识别并结合配体的活性部位;另一个是负责产生应答反应的功能活性部位。其中,负责产生应答反应的功能活性部位只有在与配体结合形成二元复合物并变构后才能产生应答反应,进而触发一系列的生化反应,最终导致靶细胞产生生物效应。受体可以诱导细胞生长、分裂和死亡,可以调控膜通道“开关”或调节细胞结合,在信号转导、免疫治疗和免疫反应中发挥重要作用。
受体-配体结合模型
1、受体特性
受体的特性是通过与配体结合的特点而产生的。受体与配体的结合是化学性的,绝对多数配体与受体是通过范德华力、离子键、氢键等分子间的吸引力来结合的,少数通过共价键结合,这样的结合较难逆转。一个真正的受体,有以下特点:
区域分布性(regional distribution):受体在生物体不同组织或同一组织的不同区域的分布密度不同。
饱和性(saturability):受体在每一细胞或每一定量组织内的数量是有限的,当配体到达一定浓度时,即使继续增加,与受体的结合值也不再改变。
特异性(specificity):特定的受体只与某种特定的配体结合,受体接合部位与配体的结构具有专一性,从而产生特定的效应。
特异性除了可以理解为一种受体仅能与一种配体结合之外,还可以表现为在同一细胞或不同类型的细胞中,同一配体可能有两种或两种以上的不同受体;同一配体与不同类型受体结合会产生不同的细胞反应,例如肾上腺素作用于皮肤粘膜血管上的α受体使血管平滑肌收缩,作用于支气管平滑肌等使其舒张。
可逆性(reversibility):配体与受体的结合是可逆的。从配体-受体复合物中解离出来的配体和受体结构不发生变化。
高亲和力(high affinity):受体对其配体的亲和力很高,相当于内源性配体的生理浓度,表观解离常数在nmol/L的水平。
内源性配体(endogenous ligand):生物体内存在受体的内源性配体,如内源性递质、激素等。
亚细胞或分子特征(subcellular or molecular characterization):同类受体不同亚型的分子量,亚细胞或分子特征各不相同。
配体结合试验资料与药理活性的相关性(binding data vs pharmacological activity relationship):受体与药物结合的强度与产生生物效应的药效强度有关。
配体结合后有生理活性:无论配体是内源性的还是药物,与受体结合后,二者形成配体-受体复合物,从而传递信号引起一系列的生理、生化效应。2、受体功能
如开篇所述,受体是细胞表面或亚细胞组分中的一种分子,可以识别并特异地与有生物活性的化学信号物质(配体)结合,从而激活或启动一系列生物化学反应,最后导致该信号物质特定的生物效应。
受体的功能通常可分为两个方面:
识别特异的信号物质--配体,识别的表现在于两者结合。
配体与受体的结合是一种分子识别过程,它靠氢键、离子键与范德华力的作用,随着两种分子空间结构互不程度增加,相互作用基团之间距离酒会缩短,作用力就会大大增加,因此分子空间结构的互补性是特异结合的主要因素。同一配体可能有两种或两种以上的不同受体,例如乙酰胆碱有烟碱型和毒蕈型两种受体,同一配体与不同类型受体结合会产生不同的细胞反应。如Ach可以使骨骼肌兴奋,但对心肌则是抑制的。
配体指的是一些除了与受体结合外本身并无其他功能的信号物质,它不能参加代谢产生有用产物,也不直接诱导任何细胞活性,更无酶的特点,它唯一的功能就是通知细胞在环境中存在一种特殊信号或刺激因素
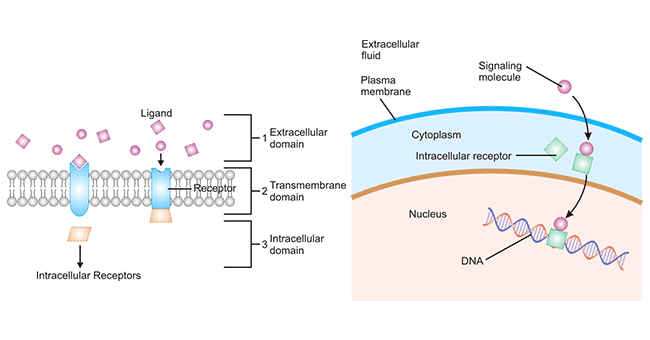
另一方面是把识别和接受的信号准确无误的放大并传递到细胞内部,激活或触发一系列胞内生化反应,最后导致特定的细胞反应。使得胞间信号转换为胞内信号。3、受体类型(图一)
目前已知的受体有30余种,按药理学和分子生物学的命名方法,按特异的内源型配体命名;
对尚不知道内源性配体的,按药物名命名。对于有亚型的受体,按受体结构类型命名,用数字和阿拉伯数字表示。
按位置分
- 细胞表面受体
- 细胞膜受体:位于靶细胞膜上,如胆碱受体、肾上腺素受体、胰岛素受体等。
- 细胞表面受体通常由三个结构域组成:细胞外配体结合结构域、嵌在质膜内的疏水结构域和细胞内结构域。
- 细胞内受体
- 胞浆受体:位于靶细胞浆内,如性激素受体、肾上腺皮质激素受体等。
- 胞核受体:位于靶细胞核内,如甲状腺素受体等。
- 细胞内受体通常位于胞质中,受体要与相应的配体结合后才可进入细胞核。胞内受体识别和结合的是能够穿过细胞质膜的小的脂溶性的信号分子,如各种类固醇激素、甲状腺素、维生素D以及视黄酸。严格来说,细胞内受体可根据受体是在胞质内还是细胞核内进一步分为胞浆受体和胞核受体。胞浆受体位于靶细胞的胞浆内,如肾上腺皮质激素受体、性激素受体。胞核受体位于靶细胞的细胞核内,如甲状腺素受体。
按特征分
根据受体的蛋白结构、信息转导过程、效应性质、受体位置等特点
-
含离子通道的受体(channel-linked receptors),又称离子通道型受体(ionotropic receptors)或离子促进型受体,是具有连接有离子通道的膜受体。根据其生理功能有可分为配体门控离子通道(ligand-gated ion channels)和电压门控离子通道(voltage-gated ion channels)。如N乙酰胆碱受体、γ-氨基丁酸受体、谷氨酸受体等。
- G蛋白偶联受体(G-protien coupled receptors)是通过G蛋白连接细胞内效应系统的膜受体。如M乙酰胆碱受体、肾上腺素受体、多巴胺受体、5-羟色胺受体、前列腺素受体等。
- 具酪氨酸激酶活性的受体(tyrosine kinase-linked receptors)是结合细胞内酪氨酸激酶范围的膜受体。如胰岛素受体、胰岛素样生长因子受体、表皮生长因子受体、血小板生长因子受体、集落刺激因子-1受体、成纤维细胞生长因子受体等。
- 调节基因表达的受体(regulate gene transcription receptors),又称核受体(nuclear receptors)、细胞内受体。如甾体激素受体、甲状腺素受体等。
图一、细胞表面受体与细胞内受体作用机制图
https://i.postimg.cc/pX4NBmFT/receptor-1.png
{kind=link}
4、受体调节
受体调节(receptor regulation)是指由于与配体作用,有关受体数目和亲合力的变化。
简介
靶细胞表面受体被其配体激动后不仅触发一系列生理生化或药理反应,
而且受体本身的数目或亲和力也发生变化。
同种或异种受体调节都是细胞反应的一部分,
它们的控制环节涉及受体蛋白的生物合成、受体的内移入胞及分解代谢或再循环、受体在脂质双层中位置的变化。
受体调节的生化机制包括受体蛋白的化学修饰以及多种因素的非共价键相互作用。
受体调节的分类
按效果分
- 向上调节(up regulation)是一种上增性调节[1],结果是受体数目增加,表现敏感性增高。通常长期使用受体的拮抗剂会产生此种效应。如长期使用普萘洛尔等药物,停药后会出现甲状腺功能亢进。
- 向下调节(down regulation)是一种衰减性调节[1],结果是受体数目下降,表现敏感性降低。通常长期使用受体的激动剂会产生此种效应。如长期使用异丙肾上腺素治疗哮喘,其疗效会不断下降。
- 在特定的生理或病理状态下,受体与其配体的结合减少(受体数目或亲和力减少)称衰减性调节,反之则称上增性调节
按种类分
- 同种调节(homospecific regulation)为配体作用于特异性地受体,使自身的受体发生变化。
- 如胰岛素受体、乙酰胆碱受体、β-肾上腺素受体、生长素受体、促甲状腺素释放激素受体、黄体生成素受体、血管紧张素Ⅱ受体等一些肽类的受体都存在同种调节作用。
- 同种调节是受体激动后的结果之一,也就是说受体激动后不仅引起靶细胞的其他反应,受体本身的数目和亲和力也发生变化
- 异种调节(heterospecific regulation)也称外调节(trans regulationb)为配体作用于其特异性的受体,对另一种配体的受体产生调节作用。
- 如维生素A可使胰岛素受体产生向下调节;血管活性肽可调节M受体;甲状腺素、糖皮质激素和性激素可调节β-肾上腺素受体;氨甲酰胆碱可调节α-肾上腺素受体;苯二氮卓可调节γ-氨基丁酸受体,苯二氮卓受体又可被γ-氨基丁酸调节;β-肾上腺素可调节胰岛素受体和表皮生长因子(EGF)受体;胰岛素样生长因子Ⅱ(IGF-Ⅱ)受体可被胰岛素调节;表皮生长因子-尿抑制素(EGF-URO)受体可被加压素、血小板生长因子和巴豆油脂肿瘤促进剂TPA所调节等。
- 异种调节也是受体激动后细胞反应的一部分
受体调节的控制环节(图二)
-
受体蛋白的生物合成 即转录、翻译、受体蛋白的糖基化及其它加工。
- 受体蛋白的生物合成受激素控制。
- 受体蛋白对包浆膜的嵌入和受体在膜脂质双层中位置的改变,在完整脂肪细胞中。
- 研究受体蛋何整合到脂肪细胞的细胞膜(胞浆膜)中,并探讨其在脂质双层中的动态位置变化,且这些研究是在保持脂肪细胞结构完整(未被破坏)的条件下进行的。以下为研究
- 在15~37°C时,胰岛素在几分钟内可增加IGF-II受体的亲和力而不影响其受体数目。
- 抗微管和抗微丝药物可取消这一效应,因此认为胰岛素对IGF-II受体的异种上增性调节,
- 可能是作用于受体对膜嵌入这一环节。
- 另一种异种调节可能是作用于受体在脂质双层中的位置。
- 例如,TPA作用于钙-磷脂依赖性蛋白激酶,间接改变受体的位置,使受体处于被“扣押”状态(seques-tration),减少EGF-URO受体的数目和/或亲和力,这种作用是快速的、温度依赖的,需要完整的细胞。
- 血小板生长因子和加压素分别作用于自已的受体上,通过类似TRA的方式,减低EGF-URO受体的亲和力。
- 受体的内移、分解代谢或再循环某些肽类配体、α-巨球蛋白以及低密度脂蛋白等的受体与配体结合后形成配体-受体复合物,通过覆衣凹陷(coatedpits)内移入胞,形成胞内体(endosome),在胞浆内贮存加工,经溶酶体降解而进行分解代谢。
- 此外,β-肾上腺素受体也可内移入胞。例如,蛙红细胞与异丙肾上腺素孵育后,其表面[3H]双氢心得舒的最大结合数Bmax下降,与此同时,胞液内[3H]双氢心得舒结合部位的Bmax上升。这种再分布过程依赖于时间和温度,需代谢能量;但不依赖于蛋白质合成。β-肾上腺素受体内移过程中,与其偶联的鸟嘌呤核苷酸调节蛋白和腺苷酸环化酶并不发生内移,刀豆球蛋白A和钙调素抑制剂可阻断β-肾上腺素受体的内移。
- 有研究表明,受体内移是可逆的,至少有部分受体内移后,不经降解而直接回到细胞表面,即受体可再循环(recycle)。目前认为,由配体触发的受体内移入胞是调节细胞表面受体密度的主要方式。通常是一种同种受体调节,但由于一种配体(特别是在高浓度时)可与它种受体产生交叉作用(crossover),因此,异种受体调节也可导致受体内移。
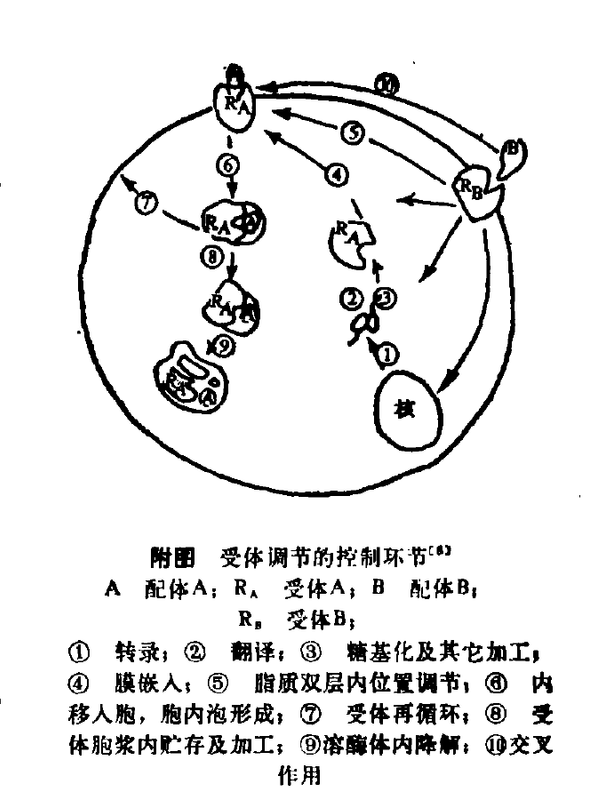{kind=link}
受体调节的生化机制
化学修饰
-
受体磷酸化与去磷酸化:这是同种及异种受体调节的机理之一。
- 光受体(视紫红质)、N胆碱受体、EGF-URO受体、胰岛素受体、血小板生长因子受体,以及β-肾上腺素受体,均有受体磷酸化。受体磷酸化有两种形式:(1)受体本身不含蛋白激酶活性,在配体直接作用下,受体的构象发生变化,成为膜结合态蛋白激酶的底物,而被磷酸化;(2)受体本身具有内在的蛋白激酶活性,在激动剂的作用下,受体的激活性得以表现,使受体自身(在ATP存在下)被磷酸化。视紫红质和N胆碱受体的磷酸化属于前一种形式,胰岛素受体、EGF-URO受体及血小板生长因子受体的磷酸化则属于后一种形式。在完整细胞,EGF-URO受体的磷酸化可发生于多个丝氨酸、苏氨酸和酪氨酸残基上。磷酸化后的EGF-URO受体可被内源性膜结合态磷酸酶水解而去磷酸化。受体可经受各种磷酸化和去磷酸化反应,受体既是磷酸基团的提供者,也是接受者。
- 硫硫键和统基互变反应:这种反应不仅对受体活性结构有重要影响,而且可能在同种受体调节中起一定作用。
- 例如,电鱼电器官上的N胆碱受体经二硫苏糖醇还原后,对琥珀酰胆碱的反应减弱,而对十烃季铵的反应增强。胎盘膜碎片经二硫苏糖醇还原后,胰岛素高亲和力结合部位消失。纯化的胰岛素受体与胰岛素结合后,受体中的基去屏蔽(unmasking),使受体从流体力学型RⅠ转变为RⅡ,同时对胰岛素的亲和力下降,这时如加入氧化型谷胱甘肽,胰岛素受体又重新转变为高亲和力的RⅠ型,因而认为硫硫键和巯基互变反应是胰岛素受体同种调节的生化机理之一。在疏基复活剂N-乙基顺丁烯二酰亚胺存在时,异丙肾上腺素的结合可导致约50%的β-肾上腺素受体识别部位失活,失活的识别部位在37℃、20min后又可自发地复活。二阶阳离子和鸟嘌岭核苷酸可促进这种复活。这种由激动剂而不能由拮抗剂结合所引起的可逆性变化与鸟呤核苷酸调节蛋白的必需基团一—巯基的变化有关。也就是说,以cAMP为细胞内信号分子的多种药物的去敏感现象,可能与上述巯基中介的机理有关。
- 受体蛋白的水解,许多激素的受体对蛋白酶敏感,但蛋白酶不一定完全取消受体的活性。
- 例如,中等浓度的蛋白酶,仅使胰岛素受体的结合曲线以及胰岛素剂量-效应曲线右移。蛋白酶可取消EGF-URO受体的蛋白激酶活性,但不取消与配体结合的能力。鉴于细胞可对多种药物刺激应答而分泌蛋白酶,而且,胞浆内的蛋白酶又可被Ca2+激活,所以,受体蛋白水解反应可能是受体调节的机理之一。
非共价键相互作用
膜电位的改变:理论上,膜电位的变化对膜上蛋白质排列的方向有极重要的影响,它参与钠离子通道等电压敏感性离子通道的调控。一种配体引起的膜电位变化完全可能导致同一细胞上其它受体结合活性的改变。
细胞表面受体分布状态的变化:细胞膜上包括受体在内的各种组份,对细胞内外信号分子应答,处于快速可逆的排列组合和相对位置的变动中,受体分子的聚集或微团形成可减弱受体的亲和力。
例如,简单的二聚体的形成就可使受体亲和力减低30%左右。这种亲和力的变化幅度虽小,但对生物系统反应性的影响却颇大。若一种受体的激活导致细胞内微管或微丝功能改变,继而引起细胞表面各种受体重新分布,则会出现异种受体调节。
除了微丝微管外,细胞膜结合态的钙调素也可能在这方面起作用。
例如,长期使用多巴胺能激动剂,纹状体突触后膜膜结合态钙调素含量下降,多巴胺结合部位的Bmax也显著下降,多巴胺受体的减敏或增敏状态取决于由胞浆膜向胞液位移的钙调素的量。
别构作用:大分子之间有别构作用,受体与脂质双层膜上其它蛋白质的相互作用可改变受体对其配体的结合能力。
例如,γ-氮基丁酸受体调变蛋白(GABA modulin)是一种膜的内在蛋白,它可减少γ-氨基丁酸受体识别部位的数目,苯二氮草受体激动后可逆转GABA modulin的上述作用,从而增大-氨基丁酸受体识别部位的数目。
小分子对大分子也有别构作用。
例如,Ca2+、Na+、CI-、GTP等小分子(离子)可通过别构作用调节受体的亲和力。CI-作用于氯离子通道而增加γ-氨基丁酸受体和苯二氮草受体分别地对其配体的亲和力。GTP不仅参与有关的受体占领与腺苷酸环化酶活性调控的偶联,而且参与胰高血糖素受体、血管紧张素II受体、α和β-肾上腺素受体、多巴胺受体、阿片受体、M胆碱受体以及前列腺素E受体等亲和力的调节。上述受体中,凡与激动剂结合后偶联腺酸环化酶活性升高者,如加入GTP,可降低其对激动剂的亲和力。因此,如果一种受体激动后,引起细胞内GTP水平升高,则可导致几种有关的受体亲和力同时减弱(异种衰减性调节)。
膜脂质环境的改变:膜脂质物理属性的改变可产生快速的同种或异种受体调节。
例如,磷脂甲基化产生磷酯酰胆碱,增高膜的流动性,使隐蔽的β-肾上腺素受体去屏蔽,导致其数目快速增多。值得注意的是,同一激动剂在不同的靶细胞可通过不同的途径或机理引起靶细胞反应性的衰减。
例如,β-肾上腺素受体激动剂引起鸭红细胞减敏的机理是使β-肾上腺素受体发生化学修饰,减弱了该受体与鸟嘌岭核苷酸调节蛋白的相互作用;同一激动剂引起蛙红细胞β-肾上腺素受体减敏的途径却使该受体内移入胞。
对药物过量的指导意义
-
大剂量用药:有些药物如肾上腺素类升压药、阿片类制剂及胰岛素等剂量过大时,可引起同种或异种受体调节。
- 长期用药:
- 长期使用激动剂,可使受体减敏,药物的剂量一效应曲线右移。
- 例如,长期用异丙肾上腺素治哮喘,在后期此药不灵,因为产生了β-肾上腺素受体的衰减性调节。
- 长期使用拮抗剂,则可使受体增敏
- 例如,长期使用心得安,如突然停药可出现肾上腺素能神经系统功能亢进症状,
- 因为肾上腺素受体产生了上增性调节。
- 配伍用药,由于有异种受体调节,因而某些表观上与某种受体无关的药物,对该受体可产生调节作用。
- 例如,磺酰脲使胰岛素受体产生上增性调节,维生素K5则使胰岛素受体产生衰减性调节。
参考文献m宋烈昌,周廷冲等.细胞表面受体的调节.生理科学进展,1987(01):24-28.
https://kns.cnki.net/KCMS/detail/detail.aspx?filename=SLKZ198701011&DBName=cjfqtotal&dbcode=cjfq
5、受体结构
蛋白质结构层次与受体功能的关系(图三)
一级结构(氨基酸序列)
- 定义:
- 多肽链中氨基酸的线性排列顺序,由基因编码决定。
- 对受体的意义:
- 决定受体与配体的结合特异性(如胰岛素受体的α亚基通过特定氨基酸残基识别胰岛素)。
- 突变可能导致功能异常:例如EGFR的L858R突变(肺癌中常见)导致激酶活性持续激活。
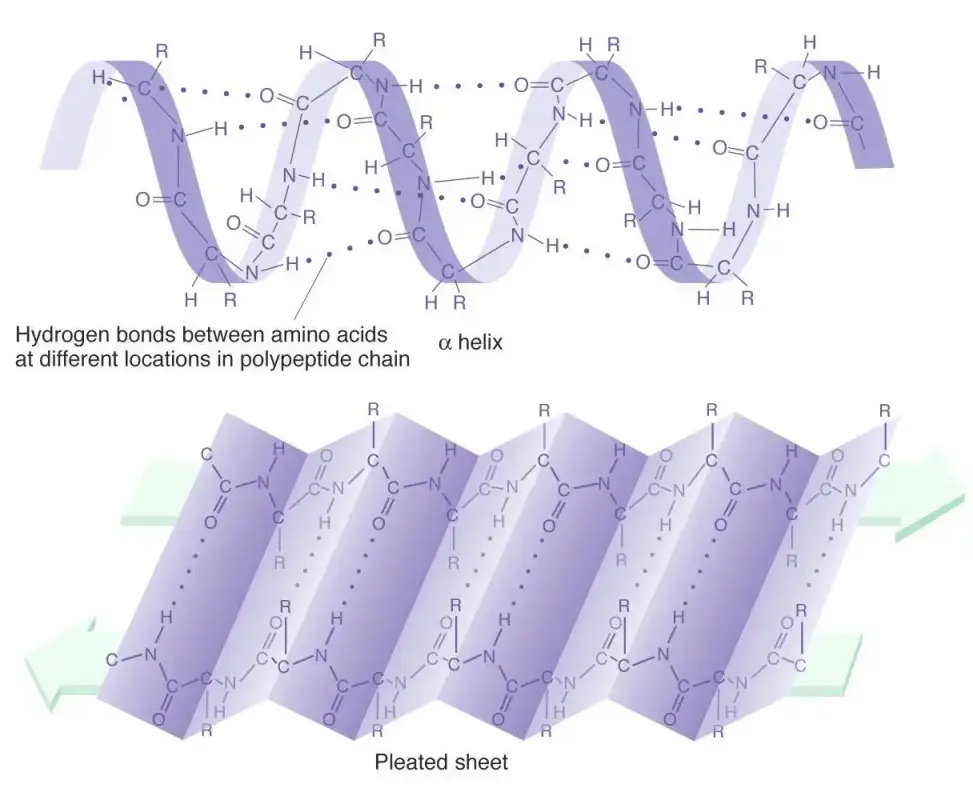
二级结构(局部折叠模式)
- 主要形式:
- α-螺旋:螺旋状构象,氢键沿螺旋轴方向形成(如跨膜受体的跨膜域)。
- β-折叠片:锯齿状链通过氢键平行或反平行排列(如抗体结合区的β桶结构)。
- 对受体的意义:
- 维持跨膜结构域的稳定性:如GPCR的7次跨膜α螺旋嵌入脂质双层。
- 参与信号传递:配体结合诱导二级结构变化(如β₂-肾上腺素受体螺旋重排暴露G蛋白结合位点)
三级结构(整体三维构象)
维持作用力:- 二硫键:半胱氨酸间共价键(如抗体铰链区的稳定性)。
- 离子键:带相反电荷侧链的静电吸引(如受体胞内域的激酶活性中心)。
- 氢键:极性基团间弱相互作用(如配体结合口袋的形成)。
- 疏水作用:非极性侧链向内聚集,避开水环境(跨膜区的疏水核心)。
对受体的意义:
- 形成配体结合位点:如雌激素受体的疏水口袋结合甾体激素。
- 调控变构效应:配体结合引发远距离结构变化(如胰岛素受体二聚化激活激酶域)。
四级结构(多亚基组装)
组成:- 多肽链亚基:如胰岛素受体为α₂β₂四聚体。
- 辅因子:非蛋白成分(辅基或辅酶),如血红素(细胞色素C受体中的铁卟啉)。
- 辅因子:非蛋白成分(辅基或辅酶),如血红素(细胞色素C受体中的铁卟啉)。 辅因子:非蛋白成分(辅基或辅酶),如血红素(细胞色素C受体中的铁卟啉)。
- 对受体的意义:
- 功能协同性:亚基间协作增强信号传递(如GABA受体由5个亚基组成氯离子通道)。
-
动态调控:亚基解离/重组调节活性(如钙调蛋白与Ca²⁺结合后构象改变)。
图三多肽链的二级结构
https://i.postimg.cc/8knNbd9L/Protein-structure.webp
{kind=link}
受体结构与功能的典型实例
G蛋白偶联受体(GPCR)结构特征:
-
七次跨膜α螺旋(二级结构)。
- 胞外N端参与配体识别,胞内环与G蛋白偶联(三级结构)。
功能机制:
配体结合引起跨膜螺旋位移,暴露G蛋白结合位点,触发信号级联。
核受体(如雌激素受体ER)
结构特征:
配体结合域(LBD)为疏水口袋(三级结构)。
DNA结合域(DBD)含锌指结构(二级结构)。
功能机制:
激素结合诱导受体二聚化(四级结构),结合DNA调控转录。 - 胞外N端参与配体识别,胞内环与G蛋白偶联(三级结构)。
结构异常与疾病关联
- 囊性纤维化:CFTR氯离子通道的F508del突变导致三级结构错误,无法定位至细胞膜。
- 癌症:HER2受体酪氨酸激酶的四级结构异常(过表达或二聚化增强)促进细胞增殖。
- 阿尔茨海默病:β-淀粉样蛋白二级结构(β-折叠)异常聚集形成神经斑块。
文章标题:受体蛋白
文章链接:https://www.overdose.day/253.html
本站所有文章除特别声明外,均采用 CC BY-NC-SA 4.0 许可协议,转载请注明来自sakura !